SUBSTANSI GENETIK
Dalam kehidupan sehari-hari pasti kita sering
mendengar kata DNA. Tes DNA yang sering kita lihat pada film-film jaman
sekarang berfungsi untuk membuktikan seorang anak benar-benar adalah anak
kandung dari orangtua tersebut. Apa yang terdapat dalam DNA sehingga bisa
digunakan dalam tes? Selain hal tersebut
ada juga rekayasa genetika dan klonging. Bagaimana hal tersebut bisa
terjadi. Penyakit-penyakit dan kelainan yang dialami seseorang juga berhubungan
dengan substansi genetik ini. Untuk dapat memahami hal-hal ini kita harus
belajar mengenai substansi genetika
1)
KROMOSOM
Bagian utama sebuah sel terdiri atas sitoplasma dan
nukleosom. Di dalam nukleus terdapat benang-benang halus yaitu kromatin. Kromatin terpintal menjadi
kromosom. Kromosom adalah suatu
struktur padat yang terdiri dai komponen molekul yaitu protein dan DNA.
Struktur padat kromosom dapat terlihat pada masa pembelahan sel pada tahap metafase.
Nukleosom (Unit Dasar Kromosom)
DNA - Histon – Nukleosom –
Selenoid – Kromatin – Kromatid – Kromosom
 -
DNA: sejenis asam nukleat yang
tergolong biomolekul utama penyusun berat kering setiap organisme
-
DNA: sejenis asam nukleat yang
tergolong biomolekul utama penyusun berat kering setiap organisme
-
Histon: kompleks dari asam
deoksiribonukleat, protein histon dan protein non histon yang ditemukan pada
inti sel eukariota.
-
Nukleosom: struktur dari kromatin
pada eukariota
-
Selenoid:
-
Kromatin:
-
Kromatid: salah satu dari dua
lengan hasil replikasi kromosom
- Kromosom: struktur di dalam sel berupa deret panjang molekul yang terdiri dari satu molekul DNA dan berbagai protein terkait yang merupakan informasi genetik suatu organisme,
b)
Bagian-Bagian dan Bentuk Kromosom
 |
i)
Kromatid: salah satu dari dua
lengan hasil replikasi kromosom. Kromatid masih melekat satu sama lain pada
bagian sentromer. Istilah lain untuk kromatid adalah kromonema. Kromonema
merupakan filamen yang sangat tipis yang terlihat selama tahap profase (dan
kadang-kadang pada tahap interfase). Kromonema sebenarnya merupakan istilah
untuk tahap awal pemintalan kromatid. Jadi, kromonema dan kromatid merupakan
dua istilah untuk struktur yang sama.
ii)
Kromomer: penebalan pada
kromonema. Kromoner merupakan struktur manik-manik yang merupakan akumulasi
dari materi kromatin yang terkadang terlihat saat interfase.
iii)
Sentromer: daerah kontriksi
disekitar pertengahan kromosom. Pada sentromer juga terdapat kinetokor yang
merupakan bagian pelekatan benang-benang spindel selama pembelahan inti dan
merupakan tempat melekatnya kromosom.
v)
Telomer: merupakan daerah terujung
pada kromosom. Telomer berfungsi untuk menjaga stabilitas bagian terujung
kromosom agar DNA di daerah tersebut tidak terurai. Maka dari itu, sel yang
telomer kromosomnya mengalami kerusakan umumnya segera mati.
- Telosentrik : sentromer terletak di ujung kromosom sehingga kromosom hanya memiliki sebuah lengan dan berbentuk seperti huruf I. Kromosom manusia tidak ada yang berbentuk telosentrik.Letak sentromer pada kromosom dapat membedakan jenisk kromosom. Berdasarkan letak sentromer, kromosom dibagi menjadi:
 |
-
Akrosentrik : sentromer terletak
di dekat ujung kromosom. Satu lengan kromosom sangat panjang, sedangkan lengan
lainnya sangat pendek
.
.
-
Submetasentrik : sentromer
terletak di submedian (ke arah salah satu ujung kromosom) dan membagi lengan
kromosom menjadi dua lengan yang tidak sama panjang. Satu lengan panjang dan
satu lengan pendek, seperti huruf L.
-
Metasentrik : sentromer terletak
di tengah, membagi lengan kromosom menjadi dua lengan yang hampir sama panjang
seperti huruf V.
c)
Tipe dan Jumlah Kromosom
Macam-macam kromosom:
-
Autosom: kromosom tubuh yang tidak
menentukan jenis kelamin. Pada manusia, pria dan wanita, bentuk dan jumlahnya
sama.
-
Genosom: kromosom seks yang
menentukan jenis kelamin. Jenis kelamin laki-laki(XY) dan perempuan(XX).
Memiliki bentuk dan jenis yang berbeda.
Kromosom dapat dibedakan antara satu dengan yang lain dari
penampilannya, hal ini karena ukuran kromosom dan posisi sntromer yang berbeda.
Kariotipe adalah tampilan visual
kromosom setiap individu.
Dalam sel somatik manusia terdapat 46 kromosom yang disusun
berpasangan mulain dari yeng paling panjang sampai paling pendek. 23 kromosom
berasal dari ayah dan 23 kromosom berasal dari ibu. Manusia memiliki 22 pasang
autosom dan 1 pasang genosom.
·
Kromososm homolog : pasangan
kromosom yang memiliki panjang, posisi sentromer dan pewarnaan yang sama
·
Lokus: posisi gen dalam kromos0m
d)
Peran Kromosom dalam Membawa Sifat
Individu
Kromosom tersusun dari DNA
dan protein
DNA merupakan molekul panjang
yang menyimpan informasi genetik. Total informasi genetik yang terdapat pada
DNA disebut genom.
Di dalam genom terdapat gen
–gen yang berisi informasi mengenai karakter tiap gennya.
2)
GEN
Gen adalah unit instruksi untuk menghasilkan atau
mempengaruhi suatu sifat herediter tertentu.
Gen terdiri dari DNA yang diselubungi atau diikat oleh
protein.
a)
Letak dan Simbol Gen
Letak suatu gen disebut lokus.
Sel diploid memiliki dua
lokus untuk setiap karakternya.
Letak gen-gen pada kromosom
disimbolkan dengan:

Simbol gen yang menunjukan
sifat resesif biasanya ditulis dengan huruf kecil
3)
ALEL

a) Letak dan Simbol AlelAlel disebut juga sebagai versi alternatif gen yang menjelaskan adanya variasi pada pewarisan suatu sifat
Fenotip: penampakan fisik
organisme, karakteristik atau ciri yang bisa dilihat (tinggi, pendek)
Genotip: penyusun genetik
organisme, susunan genetis
Homozigot
dominan : MM
Homozigot
resesif: mm
Heterozigot:
Mm
4)
DNA
DNA (deoxyribonucleic acid) atau asam deoksiribosa nuklet (ADN)
merupakan tempat penyimpanan informasi genetik.
DNA merupakan makromolekul
polinukleotida yang tersususn atas polimer neuklotida yang berulang-ulang
tersususun rangkap.
DNA membentuk heliks ganda.
Setiap nukleotida tersusun
dari :
1. Gula
deoksiribosa.
2.
Basa nitrogen =
purin : Adenin dan Guanin ( A dan G ).
Pirimidin : Sitosin dan Timin ( S/C dan T ).
A selalu berpasangan dengan T.
S/C selalu berpasangan dengan G.





3.
Gugus phosfat.
-
Setiap
gugus phospat menghubungkan
Ujung
3” karbon pada gula ke 5” karbon
Pada
gula berikutnya sepanjang rangkanya
-
Pasangan S
dan G
Memiliki 3 ikatan hidrogen
-
Pasangan A
dan T memiliki 2 ikatan hidrogen
-
Kedua
untai bergerak dari arah 3” ke 5”, dua rantai yang anti Paralel.
POLARITAS
• Polaritas terjadi karena salah satu ujung
rantai DNA merupakan gugus phospat dengan C 5”- deoksiribosa , sementara ujung
DNA lain merupakan gugus hidroksil dengan C 3”- deoksiribosa.
•
Dengan
demikian rantai polinukleutida merupakan suatu polaritas polinukleutida
3”----------5” dan 5”----------3”
•
Maka jika
digambarkan adalah sbb:
•
5”- A T T
G T S G A G G – 3”
3”- T A A S A G S T S S – 5”
•
•
Polaritas rantai polinukleotida
•
Ikatan fosfodiester yang menghubungkan molekul mononukleotida.
-
biru: gugus fosfat
-
hijau: basa nitrogen
-
kuning: ikatan fosfodiester
Ketentuan Chargaff
•
Basa
nitrogen dalam nukleotida tidak berjumlah sama rata,prosentase berbeda antara
satu spesies dengan spesies lainnya.
•
Tetapi
jumlah Adenin akan selalu sama dengan jumlah Timin dalam setiap molekul DNA.
•
Jumlah
Guanin selalu sama dengan jumlah Citosin dalam setiap molekul DNA.
b)
Replikasi DNA


5)
RNA
RNA (Ribonucleic acid) atau asam ribonukleat merupakan
makromolekul yang berfungsi sebagai penyimapanan dan penyalur informasi genetik
dan enzim (ribozin) yang dapat mengkatalis formasi RNA-nya sendiri atau molekul
RNA lain.
a)
Struktur RNA
RNA merupakan rantai tunggal
polinukleotida
Setiap ribonukleotida terdiri
dari:
-
5 karbon (ribosa)
-
Basa nitrogen
Purin : Adenine (A) dan Guanine (G)
Pirimidin: Citosin (C) dan Urasil (U)
-
Gugus fosfat


Urasil Ribosa
b)
Tipe RNA
i)
RNAd (RNA duta)/ mRNA (messenger
RNA) : membawa kode genetik ke
ribosom

ii)
RNAr (RNA ribosomal): komponen
utama ribosom
iii)
RNAt (RNA transfer): membawa asam amino ke ribosom

c)
Perbedaan antara RNA dan DNA
 |
6)
KODE GENETIK
 |
a)
Sintesis Protein
Ekspresi
gen
proses
dimana informasi yang dikode di dalam gen diterjemahkan menjadi urutan asam amino
selama sintesis protein.
i)
Transkripsi
Transkripsi merupakan urutan rantai nukleotida template
dari suatu DNA untai ganda disalin buat menghasilkan 1 rantai molekul RNA. Di inti sel.
- • Sintesis RNA dari rantai DNA (sense – antisense)
- • Unit transkripsi
- • Enzim Polymerase
- • Membutuhkan ATP
 |
- • Inisiasi: daerah DNA dimana RNA polimerase melekat dan mengawali proses transkripsi.
- • Elongasi: Waktu RNA bergerak di sepanjang DNA, pilinan heliks ganda DNA terbuka secara berurutan. Enzim RNA polimerase menambahkan nukleotida ke ujung dari molekul RNA yang sedang tumbuh. Setelah sintesis, DNA heliks ganda terbentuk kembali dan molekul RNA baru lepas dari cetakan DNA – nya.
- • Terminasi: suatu urutan DNA untuk menghentikan proses transkripsi.
ii)
Translasi
- Translasi merupakan sintesis polipeptida dengan urutan spesifik berdasarkan rantai RNA.
- • Inisasi: subunit ribosom kecil mengikatkan diri pada RNAd dan RNAt inisiator. Sub-unit ibosom kecil melekat pada tempat tertentu di ujung dari RNAd. Di tempat pelekatan ribosom sub-unit kecil pada RNAd ada kodon AUG, untuk memberikan sinyal dimulainya proses. RNAt inisiator, membawa asam amino metionim dan melekat di kodon inisiasi AUG.
- • Elongasi: asam amino ditambahkan 1/1 pada asam amino pertama, kodon RNAd pada ribosom membentuk ikatan hidrogen dengan anti kodon molekul RNAt yang komplimen.
- • Terminasi: Ribosom mencapai kodon terminasi pada untai RNAd. Polipeptida lepas dari ribosom, polipeptida bebas. Dua sub unit ribosom dan komponen lainnya terpisah-pisah.
- • Sel menginterpretasikan kode genetik à protein yang sesuai.
- • Kodon RNAd >< antikodon RNAt
- • Aminoasil-RNAt sintetase



Substansi Genetik
Translasi
Transkripsi
SOAL DAN PEMBAHASAN
1. Gen adalah subtansi genetika yang membawa sifat menurun. Gen terletak pada …
a. Nucleus
b. Lokus
c. Kromosom
d. Sitoplasma
e. Protoplasma (Ebtanas ’86)
kunci: B
2. Pada gambar terlihar sebagian model DNA dan replikasi DNA. Rantai tangganya mulai terpisah pada suatu bagian ujung dan nukleotida bebas mulai bergabung dengan rantai tersebut. Jika senyawa 1, 2 dan 3 adalah guanine, sitosin dan timin, maka bagian 5, 6 dan 7 adalah …
a. Sitosin, guanine, adenine
b. Sitosin, guanine, timin
c. Timin, adenine, guanine
d. Adenine, timin, guanine
e. Sitosin, timin, adenine (Ebtanas ’86)
Kunci: A
3. Kromosom yang membawa sifat menurun dari organisme terletak di …
a. Sitoplasma
b. Nucleus
c. Nucleolus
d. Plastid
e. Mitokondria (Ebtanas ’87)
kunci: C
1. Gen adalah subtansi genetika yang membawa sifat menurun. Gen terletak pada …
a. Nucleus
b. Lokus
c. Kromosom
d. Sitoplasma
e. Protoplasma (Ebtanas ’86)
kunci: B
2. Pada gambar terlihar sebagian model DNA dan replikasi DNA. Rantai tangganya mulai terpisah pada suatu bagian ujung dan nukleotida bebas mulai bergabung dengan rantai tersebut. Jika senyawa 1, 2 dan 3 adalah guanine, sitosin dan timin, maka bagian 5, 6 dan 7 adalah …
a. Sitosin, guanine, adenine
b. Sitosin, guanine, timin
c. Timin, adenine, guanine
d. Adenine, timin, guanine
e. Sitosin, timin, adenine (Ebtanas ’86)
Kunci: A
3. Kromosom yang membawa sifat menurun dari organisme terletak di …
a. Sitoplasma
b. Nucleus
c. Nucleolus
d. Plastid
e. Mitokondria (Ebtanas ’87)
kunci: C
{kind=link}
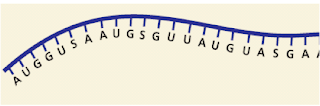
5. Dibawah ini sepotong molekul DNA yang belum lengkap pasangan basanya

Basa nitrogen yang diberi nomor adalah …
a. GUAS
b. ATAT
c. GTAS
d. ATUS
e. GUAT (Ebtanas ’87)
kunci: C
6. Berikut basa nitrogen yang menyusun DNA
1. Adenin
2. Sitosin
3. Guanine
4. Timin
Basa nitrogen dari golongan pirimidin adalah …
a. 1 dan 2
b. 2 dan 3
c. 3 dan 4
d. 2 dan 4
e. 1 dan 3 (Ebtanas ’88)
kunci: D
7. Gambar kromosom dibawah ini adalah anatomi kromosom. Bagian kromosom yang berlabel 3 dan 5 adalah …

a. Sentromer dan kromonemata
b. Matriks dan selaput kromosom
c. Selaput kromosom dan kromonemata
d. Sentromer dan matriks
e. Kromonemata dan lengan kromosom (Ebtanas ’89)
8. Pergunakan table kodon untuk menjawab soal ini!

Kodon Asam amino
Jika RNAd yang ditranskripsi DNA seperti rantai di bawah ini, maka rangkaian asam amino yang terbentuk adalah …
AAC GUG CGA UGU
A. Valin – lisin – alanin – sistein
B. Lisin – alanin – sistein – prolin – valin
C. Lisin – valin – prolin – sistein
D. Lisin – valin – prolin – sistein
E. Prolin – alanin – lisin – sistein (Ebtanas ’89)
Kunci: C/D
9. Salah satu fungsi DNA adalah menyusun protein. Sintesis ini terjadi di dalam ribosom. Mekanisme sintesa protein ini berturut-turut dari asam nukleat, DNA dan RNA dapat dinyatakan sebagai berikut:
a. DNA – RNAt – RNAd
b. RNAd – DNA – RNAt
c. DNA – RNAd – RNAt
d. RNAt – RNAd – DNA
e. RNAd – RNAt – DNA (Ebtanas ’89)
kunci: C
10. Urutan basa dalam rantai DNA membentuk kode genetic tertentu. Jika urutan basa-basa nitrogen dalam rantai nukleotida DNA adalah SSA TTA SAS STT AAT TAS, maka urutan basa nitrogen dalam rantai nukleotida DNA pelengkapnya adalah …
a. SST AAA SAS STT AAT TAS
b. GGT TAA SAS SAA TAS ATG
c. GGT AAT GAG GAA TTA ATG
d. GGT AAT GTG GAA TTA ATG
e. GGU AAU GUG GAA UUA AUG (Ebtanas ’89)
kunci: D
11. Dari rangkaian DNA dibawah ini yang dimaksud dengan symbol bernomor 1 – 2 – 3 – 4 adalah …

a. Gula – pospat – timin – guanine
b. Pospat – gula – adenine – sitosin
c. Gula – pospat – adenine – sitosin
d. Pospat – gula – timin – guanine
e. Pospat – gula – sitosin - adenin (Ebtanas '89)
Kunci: B
12. Urutan basa Nitrogen rantai sense : ATS, GGA, SST, AAG, AAS maka rantai dari ARN
duta yang terbentuk adalah ......
A . ASU, GGA, SSU, AAG, AAS
B . AAG, GGU, SSU, AAG, AAS
C . GAU, SSS, AAG, SSU, GGU
D . UGA, SUS, GAG, SUU, UGU
E . UAG, SSU, GGA, UUS, UUG
Kunci : A
Penyelesaian :
Jadi, ATS, GGA, SST, AAG, AAS berubah atau berbentuk menjadi ASU, GGA, SSU,
AAG, AAS
13 . Fungsi ADN adalah .......
A . berhubungan dengan sintesis protein dan kadarnya tetap
B . tidak berhubungan dengan sintesis protein dan kadarnya protein
C . berhubungan dengan pengendalian faktor keturunan dan sintesis protein
D . berhubungan dengan sintesis protein dan kadarnya berubah-ubah
E . berhubungan dengan sintesis protein dan faktor keturunan serta kadarnya
berubah-ubah
Kunci : A
Penyelesaian :
Fungsi ADN adalah :
- berhubungan dengan sintesis protein
- adanya tidak dipengaruhi oleh sintesis protein
- berhubungan dengan penurunan sifat.
14 . Di bawah ini tahap-tahap dari sintesis protein.
1. ARN-d melekat pada ribosom
2. ARN-d keluar dari inti masuk sitoplasma
3. ARN-t yang membawa asam amino yang sesuai
4. ARN-d disintesis oleh ADN di dalam inti
5. Asam-asam amino akan berjajar dalam urutan yang sesuai
Urutan yang benar dari tahapan sintesis protein adalah ........
A . 1 - 2 - 3 - 4 dan 5
B . 2 - 3 - 4 - 1 dan 5
C . 2 - 3 - 4 - 1 dan 5
D . 4 - 2 - 1 - 3 dan 5
E . 1 - 4 - 2 - 3 dan 5
Kunci : D
Penyelesaian :
ARNd dicetak ADN -----> ARNd keluar dari inti -----> ARNd melekat pada ribosim ----->
ARNt membawa asam amino yang sesuai ----> asam amino akan berjajar dalam
urutan yang sesuai.
15 . Bentuk kromosom disebut akrosentik bila letak sentromer .......
A . ditengah-tengah
B . agak jauh dari ujung
C . mendekati ujung
D . paling ujung
E . di kedua ujung
Kunci : C
Penyelesaian :
Jika letak kromosom :
- di tengah-tengah : metasentrik
- agak jauh dari ujung : submetasentrik
- di ujung : telosentrik
- mendekati ujung : akrosentrik